


 de INMUNOLOGÍA
de INMUNOLOGÍA

 Michael W. Russell
Michael W. Russell  Zina Moldoveanu
Zina Moldoveanu  Pearay L. Ogra 3 y
Pearay L. Ogra 3 y  Jiri Mestecky
Jiri Mestecky - 1 Departamento de Microbiología e Inmunología, Escuela Jacobs de Medicina y Ciencias Biomédicas, Universidad de Buffalo, Buffalo, NY, Estados Unidos
- 2 Departamento de Microbiología, Universidad de Alabama en Birmingham, Birmingham, AL, Estados Unidos
- 3 División de Enfermedades Infecciosas, Departamento de Pediatría, Facultad de Medicina y Ciencias Biomédicas Jacobs, Universidad de Buffalo, Buffalo, NY, Estados Unidos
El sistema inmunológico de las mucosas es el componente más grande de todo el sistema inmunológico, habiendo evolucionado para brindar protección en los principales sitios de amenaza infecciosa: las mucosas. Como el SARS-CoV-2 infecta inicialmente el tracto respiratorio superior, sus primeras interacciones con el sistema inmunológico deben ocurrir predominantemente en las superficies de la mucosa respiratoria, durante las fases inductiva y efectora de la respuesta. Sin embargo, casi todos los estudios de la respuesta inmune en COVID-19 se han centrado exclusivamente en los anticuerpos séricos y la inmunidad sistémica mediada por células, incluidas las respuestas innatas. Este artículo propone que existe un papel importante para la inmunidad de la mucosa y para los anticuerpos IgA secretores y circulantes en COVID-19, y que es importante dilucidar esto para comprender especialmente los estados asintomáticos y leves de la infección. que parecen representar la mayoría de los casos. Además, es posible que se pueda aprovechar la inmunidad de las mucosas con fines de diagnóstico, terapéuticos o profilácticos beneficiosos.
Introducción
Aunque la pandemia de COVID-19 ha estado en curso durante varios meses, se ha prestado muy poca atención a la inmunidad de las mucosas en la infección por SARS-CoV-2. Sin embargo, este virus infecta principalmente las superficies mucosas del tracto respiratorio (y posiblemente también el tracto digestivo) al menos hasta las etapas avanzadas de la enfermedad, cuando el ARN viral puede volverse detectable en la circulación ( 1). El virus también puede adquirirse a través de la boca y en la superficie conjuntival del ojo, de donde desemboca en los conductos nasales a través del conducto lagrimal. Esto significa que sus interacciones con el sistema inmunológico, durante las fases inductiva y efectora, deben ocurrir primero predominantemente, si no exclusivamente, en las mucosas respiratoria y oral. Esto tiene profundas implicaciones para los resultados y debería orientar nuestro enfoque para investigar y comprender la inmunidad adaptativa en la enfermedad COVID-19, incluido su diagnóstico, tratamiento y desarrollo efectivo de vacunas. En términos tanto del despliegue de células inmunes como de la producción de inmunoglobulinas, el sistema inmunológico de las mucosas es, con mucho, el componente más grande de todo el sistema inmunológico, habiendo evolucionado para brindar protección en los principales sitios de amenaza infecciosa: las mucosas (2 ). La IgA secretora (SIgA) se produce en cantidades muy superiores a las de todos los demás isotipos de inmunoglobulina combinados ( 3 ). Junto con el suero homólogo, que se deriva de una fuente distinta, la médula ósea, la IgA es el más heterogéneo de los isotipos de inmunoglobulina, y se presenta en tres formas moleculares (secretora, polimérica y monomérica), dos subclases (IgA1 e IgA2), y numerosas glicoformas ( 4 ), indicando colectivamente marcadas diferencias en la función fisiológica relacionadas en parte con los lugares en los que ocurren. Mientras que la IgA circulante es principalmente monomérica y consiste predominantemente en la subclase IgA1, SIgA es dimérica y consiste en proporciones variables de IgA1 e IgA2 ( Tabla 1). Se han atribuido pocas diferencias funcionales a las subclases de IgA, aparte de que son inducidas preferentemente por antígenos de proteínas frente a antígenos de carbohidratos ( Tabla 1 ), y la bisagra más larga de IgA1 le confiere una mayor flexibilidad para alcanzar epítopos antigénicos separados. Sin embargo, se han atribuido diferentes funciones efectoras de las subclases de IgA a sus diferentes perfiles de glicosilación ( 5 ).
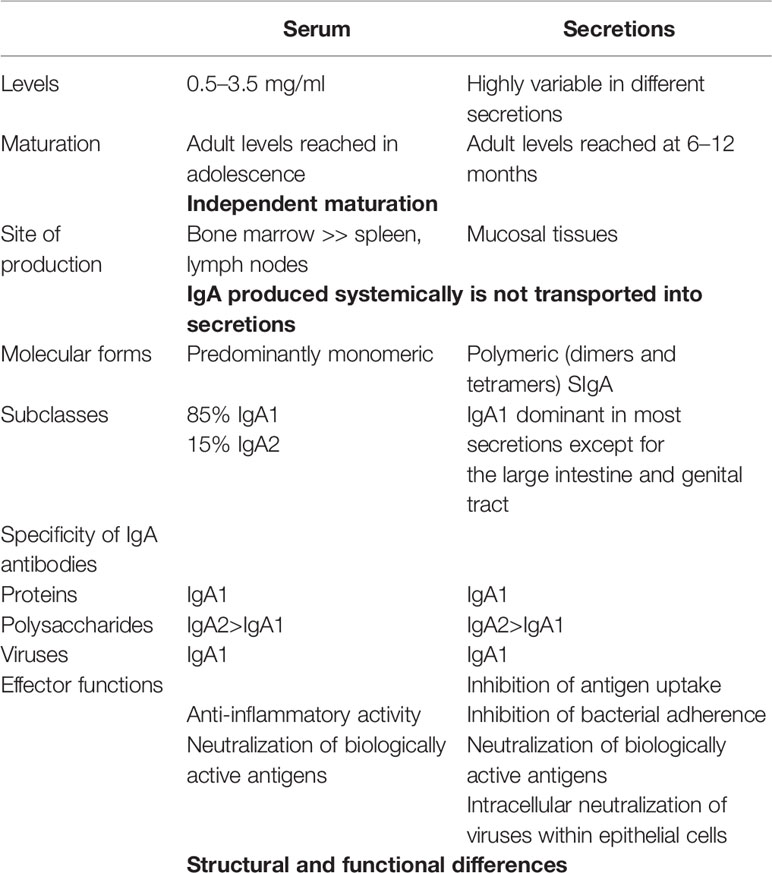 Tabla 1 Independencia de los compartimentos de IgA sistémicos y mucosos.
Tabla 1 Independencia de los compartimentos de IgA sistémicos y mucosos.
El papel de las respuestas inmunitarias de las mucosas
Dado que el SARS-CoV-2 primero infecta principalmente el tracto respiratorio superior (URT), se espera que se induzcan respuestas inmunitarias de la mucosa en la nasofaringe, tanto a través del epitelio nasal como a través de las amígdalas y adenoides, que se denominan colectivamente asociadas a nasofaringe. tejido linfoide (NALT) que sirven como sitios inductivos para el sistema inmunológico de la mucosa ( 6 , 7 ). Es posible que las respuestas también se puedan inducir a través de sitios inductivos de la mucosa en el conducto lagrimal ( 8 ) o la cavidad oral ( 9), aunque la contribución cuantitativa de dichos sitios a las respuestas inmunitarias de las mucosas en humanos es incierta. El tejido linfoide asociado a los bronquios (BALT) normalmente no está presente en los seres humanos adultos, pero se puede encontrar en niños y adolescentes, y puede inducirse a su formación por infecciones ( 10). Esto plantea preguntas interesantes sobre si las respuestas inducidas en BALT podrían contribuir a la mayor resistencia reportada de los jóvenes a la enfermedad COVID-19, o si BALT podría ser inducida por SARS-CoV-2 con consecuencias para el curso de la infección. Todos estos tejidos del sitio inductivo de la mucosa generan células B de la mucosa productoras de IgA que albergan varios sitios efectores de la mucosa remotos donde se diferencian en células plasmáticas secretoras de IgA poliméricas (p). Además, las células B productoras de IgG sistémicas también se inducen en las amígdalas y estas albergan tejidos linfoides periféricos donde se diferencian y secretan IgG para la circulación ( 11). En los espacios subepiteliales de las mucosas y las glándulas asociadas, las células plasmáticas de la mucosa producen pIgA que se transporta selectivamente a las secreciones por la vía mediada por el receptor de inmunoglobulina polimérica, que se libera como SIgA ( 12 ). Tanto en los conductos nasales como cuando desciende a la tráquea y los bronquios, el virus se encuentra con un entorno dominado por SIgA, que se genera a través del sistema inmunitario de la mucosa y mantiene un medio esencialmente no inflamatorio. Sin embargo, una vez que alcanza las vías respiratorias terminales y los alvéolos, ingresa a un ambiente dominado por IgG derivada de la circulación.
Se ha prestado mayor atención a los anticuerpos neutralizantes de virus, especialmente a los anticuerpos circulantes ( 13 – 15 ). Sin embargo, estos solo pueden ser efectivos en la prevención de infecciones o enfermedades si alcanzan las superficies mucosas donde está presente el virus, y debe tenerse en cuenta que la IgA circulante, incluso en forma polimérica, no se transporta de manera efectiva a las secreciones ( 16). Mientras que la IgG derivada de plasma se produce en la URT y especialmente en el tracto respiratorio inferior (LRT), la IgG es inflamatoria en su modo de acción, por la inducción de mecanismos efectores como la activación del complemento y la participación de fagocitos como macrófagos y neutrófilos. como células asesinas naturales (NK). La patología grave de COVID-19 ocurre en las vías respiratorias terminales de los pulmones, donde la IgG circulante es la inmunoglobulina dominante. La intensa inflamación resultante involucra múltiples factores moleculares y celulares, incluidas las células reclutadas por quimioatrayentes inducidos por virus ( 17 ). El brazo celular de la respuesta inmune adaptativa, incluidas las células T CD4 + y CD8 + citotóxicas, también se administra a través dela circulación y puede llegar a los alvéolos. Sin embargo, las células citotóxicas por su naturaleza no pueden prevenir la infección: destruyen las células ya infectadas y, por lo tanto, reducen la propagación de la infección.
Casi todos los esfuerzos en el desarrollo de vacunas contra COVID-19 se centran en la inyección sistémica, que induce predominantemente anticuerpos IgG circulatorios y, potencialmente, células T citotóxicas ( 18 ). Estas rutas son poco efectivas para generar respuestas inmunitarias de las mucosas, que solo pueden ser inducidas por las rutas de inmunización de las mucosas, incluso a través de la NALT en la URT. Las respuestas inmunitarias de las mucosas están parcialmente compartimentadas, ya que la distribución de las respuestas depende de la ruta real de inducción ( 7 , 19 ). Por ejemplo, la vía entérica genera predominantemente respuestas en el tracto gastrointestinal, mientras que la vía nasal genera predominantemente respuestas en el tracto respiratorio y las glándulas salivales ( 7). Las razones de estas distribuciones diferenciales radican en la impronta de las células T y B inducidas en los respectivos sitios inductivos, los tejidos linfoides asociados al intestino (GALT, como las placas de Peyer intestinales) o NALT, con receptores «homing» que incluyen integrinas específicas. y receptores de quimiocinas específicos para los tejidos diana ( 20). En términos prácticos, esto significa que la inmunización intranasal debería ser un medio eficaz de generar respuestas de anticuerpos predominantemente SIgA en el URT y LRT, donde el SARS-CoV-2 podría neutralizarse y eliminarse sin consecuencias inflamatorias. Además, implica que analizar los anticuerpos IgA en las secreciones nasales o la saliva debería ser una forma más informativa de evaluar las respuestas inmunitarias eficaces contra el SARS-CoV-2, ya sea inducido por la infección natural o por inmunización intranasal. El ensayo de anticuerpos IgA en suero, aunque es de interés adicional, no esun sustituto, porque la IgA sérica proviene de una fuente diferente (principalmente la médula ósea) y consiste principalmente en IgA1 monomérica. Esto es distinto de la SIgA mucosal, que consta de ambas subclases y es sintetizada localmente por células plasmáticas secretoras de pIgA que residen en los espacios subepiteliales (lámina propia) de los tejidos y glándulas mucosas ( 21 ).
Se sabe que los anticuerpos SIgA son eficaces contra diversos patógenos, incluidos los virus, mediante mecanismos como la neutralización, la inhibición de la adherencia y la invasión de las células epiteliales, la aglutinación y la facilitación de la eliminación en la corriente de moco ( 22 ). También se han descrito mecanismos intracelulares de inhibición de la replicación viral ( 23 ). Además, SIgA es esencialmente no inflamatorio, incluso antiinflamatorio, en su modo de acción. La IgA no activa el complemento por la vía clásica y la activación de la vía alternativa por la IgA es en gran medida un artefacto, mientras que la vía de la lectina depende de los residuos de azúcar terminales en las estructuras de glicanos ( 22). Además, incluso se ha demostrado que los anticuerpos IgA inhiben la activación del complemento mediada por anticuerpos IgM o IgG ( 24 , 25 ). Curiosamente, los resultados de un ensayo de inmunización sistémica contra el VIH en humanos sugieren que los niveles altos de respuestas de IgA en suero comprometieron la función protectora de los anticuerpos IgG con la misma especificidad antigénica y se asociaron con un mayor riesgo de infección por VIH ( 26 ).
Cabe señalar que mientras que los niveles de SIgA en las mucosas aumentan rápidamente en los bebés y alcanzan los niveles de los adultos en la infancia, los niveles de IgA en suero maduran mucho más lentamente y pueden no alcanzar los niveles completos de los adultos hasta la adolescencia ( 27 ). Dada la aparente diferencia en la susceptibilidad a la enfermedad por COVID-19 entre niños pequeños y adultos ( 28 ), estas diferencias en la maduración de la respuesta inmune deben considerarse.
La deficiencia selectiva de IgA es relativamente común entre las personas de ascendencia europea, donde la prevalencia puede llegar a 1 de cada 400 personas aproximadamente ( 29). La deficiencia afecta tanto a los compartimentos circulatorio como a la mucosa y los sujetos a menudo muestran una mayor susceptibilidad a las infecciones por URT. Se desconoce por completo si la deficiencia de IgA afecta a COVID-19 y cómo. Por un lado, si los anticuerpos SIgA mucosos en la URT ejercen un efecto protector contra las primeras etapas de la infección por SARS-CoV-2, se esperaría que la deficiencia de SIgA intensifique la infección, facilitando el descenso a la LRT y conduciendo a una enfermedad avanzada. Alternativamente, la ausencia de IgA circulatoria con su capacidad para interferir con los mecanismos de defensa mediados por IgM e IgG podría resultar beneficiosa en la infección por LRT, donde predominan los anticuerpos IgG. Una vez más, la ausencia de control antiinflamatorio por parte de los anticuerpos IgA podría facilitar la inflamación desregulada.
Discusión
Sostenemos que la inmunidad de las mucosas tiene un papel importante que desempeñar en COVID-19, en varios niveles. Dado que el SARS-CoV-2 primero infecta principalmente a través de las fosas nasales, posiblemente a través de los ojos, seguido de un drenaje hacia la URT, también a través de la boca, predeciríamos que las primeras respuestas inmunes deberían revelarse a través del sistema inmunológico de las mucosas, con la aparición de anticuerpos SIgA en secreciones de URT, y también en saliva y líquido lagrimal. También puede producirse la producción concomitante de anticuerpos IgG e IgA séricos ( 11). Antes de la producción de anticuerpos SIgA, debe haber una ola de células secretoras de anticuerpos IgA en la circulación. Esto ocurre típicamente con un pico alrededor de 6 a 10 días después de un evento de inmunización de la mucosa discreto, ya que las células B inducidas en los sitios inmunoinductores de la mucosa (como NALT) expresan receptores de localización mucosa como la integrina α4β7 y migran a los sitios efectores donde se diferencian terminalmente en células plasmáticas secretoras de pIgA ( 11 , 30). Las células secretoras de anticuerpos IgG circulantes también son inducidas por antígenos que estimulan respuestas en las amígdalas, y estas suelen expresar receptores de localización periféricos, como L-selectina. El período en el que se pueden detectar estas células es limitado, ya que esta ola de células B migratorias es transitoria después de un evento inductor, que en el caso de la infección por SARS-CoV-2 podría ser de hasta 4-5 días (o más) antes. los primeros síntomas aparecen. Si es así, el pico de migración celular podría ocurrir entre 4 y 5 días después de que aparezcan los síntomas. Sin embargo, en presencia de infección continua y estimulación inmune continua, es posible que las células secretoras de anticuerpos continúen circulando a medida que se liberan ondas repetidas de células B inducidas desde los sitios inductivos, como se ha informado para las infecciones por VIH ( 31). Debido a las incertidumbres en la cinética del desarrollo de anticuerpos IgM, IgG e IgA en individuos infectados, el análisis detallado de las células secretoras de anticuerpos y su expresión de receptores autodirigidos, y su persistencia en los tejidos linfoides, proporcionará información adicional esencial y más completa sobre la respuesta inmune al SARS-CoV-2. Dichos análisis podrían arrojar luz sobre las diferencias en el resultado clínico de la infección en los niños en comparación con los adultos sintomáticos o asintomáticos. Se puede esperar que la determinación de estas respuestas celulares y de anticuerpos del sistema inmunológico de la mucosa proporcione información valiosa que sea distinta y complementaria a la determinación de las respuestas de anticuerpos IgG en suero. Además, es razonable esperar que la información arroje información valiosa sobre el progreso de la enfermedad COVID-19,32 ). Sin embargo, un requisito clave para obtener resultados confiables es que los procedimientos utilizados para la recolección de muestras, así como los métodos de ensayo, deben diseñarse teniendo en cuenta las características de las respuestas inmunitarias de las mucosas y las características distintivas de las secreciones en comparación con el suero ( 33 ).
Cabe señalar que se han registrado respuestas de anticuerpos IgA en los fluidos nasales de voluntarios infectados con el coronavirus 229E del resfriado común y se han asociado con períodos más cortos de diseminación viral ( 34 ). Actualmente se han informado respuestas de anticuerpos IgA en suero y saliva a antígenos de pico del SARS-Cov-2 ( 35 ), y los anticuerpos IgA en saliva persistieron durante al menos 3 meses. Curiosamente, si bien se encontraron buenas correlaciones entre los niveles de anticuerpos IgM e IgG en suero y saliva, hubo una correlación mucho más débil entre los anticuerpos IgA en suero y salival, como se esperaba dado que la IgM e IgG salivales se derivan en gran medida de la circulación, mientras que la IgA salival se genera principalmente localmente en las glándulas salivales como SIgA ( 19). Dos preimpresiones recientes informan que los anticuerpos IgA contra el SARS-CoV-2 estaban elevados en los fluidos nasales, las lágrimas y la saliva de los sujetos infectados ( 36 , 37 ), y que los plasmablastos secretores de IgA que expresan el receptor de quimiocinas mucosas CCR10 estaban elevados en la sangre periférica de sujetos infectados con SARS-CoV-2 ( 37 ). Estos hallazgos apoyan el concepto de que las respuestas de anticuerpos IgA de las mucosas son inducidas por el SARS-CoV-2. Se esperan con gran interés nuevos estudios destinados a relacionar estas respuestas con el curso de la infección en sujetos de diferentes grupos de edad y con diferentes resultados de la enfermedad.
Finalmente, esperamos que los esfuerzos en el desarrollo de vacunas dirigidos a inducir respuestas inmunes de las mucosas y células de memoria, especialmente en la URT, produzcan beneficios que no se ven con las rutas parenterales convencionales de administración de vacunas. Las vacunas intranasales ya están disponibles contra la influenza y otras están en desarrollo ( 30 , 38 ). Las ventajas, además de la administración sin aguja, incluyen la generación de anticuerpos tanto mucosos (SIgA) como circulantes (IgG e IgA), así como respuestas de células T. Como se discutió anteriormente, tales respuestas podrían lograr resultados deseables que no se obtienen con rutas de inmunización sistémica.
En resumen, con base en la ruta por la cual se adquiere la infección por SARS-CoV-2 y la independencia de las respuestas mucosas y sistémicas, debe haber una dimensión inmunitaria mucosa al COVID-19. Si hace una contribución significativa al resultado de la infección por SARS-CoV-2 o si se puede aprovechar con buenos resultados para fines de diagnóstico o para terapia y profilaxis, solo se puede determinar mediante la realización de las investigaciones adecuadas. La Tabla 2 enumera algunos estudios potencialmente importantes que deben realizarse para dilucidar las respuestas inmunitarias de las mucosas en la infección por SARS-CoV-2.
 Tabla 2 Estudios futuros deseables sobre las respuestas inmunitarias de las mucosas al SARS-CoV-2.
Tabla 2 Estudios futuros deseables sobre las respuestas inmunitarias de las mucosas al SARS-CoV-2.
Declaración de disponibilidad de datos
Cualquier contribución original presentada en el estudio se incluye en el material del artículo.
Contribuciones de autor
Los objetivos de este artículo fueron concebidos por todos los autores. MR redactó el primer borrador, que fue revisado y enmendado por todos los autores para crear la versión final. Todos los autores contribuyeron al artículo y aprobaron la versión enviada.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Referencias
1. Bermejo-Martin JF, Gonzalez-Rivera M, Almansa R, Micheloud D, Dominguez-Gil M, Resino S, et al. La viremia del ARN del SARS-CoV-2 se asocia con una respuesta del huésped similar a la sepsis y una enfermedad crítica en COVID-19. medRxiv (2020) 2020.08.25.20154252. doi: 10.1101 / 2020.08.25.20154252
2. Russell MW, Mestecky J, Strober W, Kelsall BL, Lambrecht BN, Cheroutre H. El sistema inmunológico de las mucosas: descripción general. En: Mestecky J, Strober W, Russell MW, Kelsall BL, Cheroutre H, Lambrecht BN, editores. Inmunología de las mucosas , 4. Amsterdam: Academic Press / Elsevier (2015). pag. 3-8.
3. Russell MW. Funciones biológicas de IgA. En: Kaetzel CS, editor. Mucosa Defensa inmunológica: Inmunoglobulina A . Nueva York: Springer (2007). pag. 144–72.
4. Woof JM, Mestecky J. Inmunoglobulinas mucosas. En: Mestecky J, Strober W, Russell MW, Kelsall BL, Cheroutre H, Lambrecht BN, editores. En: Inmunología de las mucosas , 4. Amsterdam: Academic Press / Elsevier (2015). pag. 287–324.
5. Steffen U, Koeleman CA, Sokolova MV, Bang H, Kleyer A, Rech J, et al. Las subclases de IgA tienen diferentes funciones efectoras asociadas con distintos perfiles de glicosilación. Nat Communs (2020) 11: 120. doi: 10.1038 / s41467-019-13992-8
6. Brandtzaeg P. Inmunobiología de las amígdalas y adenoides. En: Mestecky J, Strober W, Russell MW, Kelsall BL, Cheroutre H, Lambrecht BN, editores. Inmunología de las mucosas , 4. Amsterdam: Academic Press / Elsevier (2015). pag. 1985-2016.
7. Boyaka PN, McGhee JR, Czerkinsky C, Mestecky J. Vacunas mucosas: una descripción general. En: Mestecky J, Bienenstock J, Lamm ME, Mayer L, Strober W, McGhee JR, editores. Inmunología de las mucosas , 3. Amsterdam: Academic Press / Elsevier (2005). pag. 855–86.
8. O’Sullivan NL, Montgomery PC. Inmunidad de la mucosa ocular. En: Mestecky J, Strober W, Russell MW, Kelsall BL, Cheroutre H, Lambrecht BN, editores. Inmunología de las mucosas , 4. Amsterdam: Academic Press / Elsevier (2015). pag. 1873–97.
9. Czerkinsky C, Cuburu N, Kweon MN, Anjuere F, Holmgren J. Vacunación sublingual. Hum Vacc (2011) 7: 110–4. doi: 10.4161 / hv.7.1.13739
10. Tschernig T, Pabst R. El tejido linfoide asociado a bronquios (BALT) no está presente en el pulmón adulto normal sino en diferentes enfermedades. Pathobiology (2000) 68: 1–8. doi: 10.1159 / 000028109
PubMed Abstract | CrossRef Texto completo | Google Académico
11. Quiding-Järbrink M, Nordström I, Granström G, Kilander A, Jertborn M, Butcher EC, et al. Expresión diferencial de moléculas de adhesión específicas de tejido en células formadoras de anticuerpos circulantes humanos después de inmunizaciones sistémicas, entéricas y nasales. Una base molecular para la compartimentación de las respuestas de las células B efectoras. J Clin Invest (1997) 99: 1281–6. doi: 10.1172 / JCI119286
PubMed Abstract | CrossRef Texto completo | Google Académico
12. Baker K, Blumberg RS, Kaetzel CS. Transporte de inmunoglobulinas y receptores de inmunoglobulinas. En: Mestecky J, Strober W, Russell MW, Kelsall BL, Cheroutre H, Lambrecht BN, editores. Inmunología de las mucosas , 4. Amsterdam: Academic Press / Elsevier (2015). pag. 349–407.
13. Klingler J, Weiss S, Itri V, Liu X, Oguntuyo KY, Stevens C, et al. Papel de los anticuerpos IgM e IgA en la neutralización del SARS-CoV-2. medRxiv (2020) 2020.08.18.20177303. doi: 10.1101 / 2020.08.18.20177303
PubMed Abstract | CrossRef Texto completo | Google Académico
14. Ejemel M, Li Q, Hou S, Schiller ZA, Tree JA, Wallace A, et al. Un anticuerpo monoclonal IgA humano de reacción cruzada bloquea la interacción SARS-CoV-2 pico-ACE2. Nat Communs (2020) 11: 4198. doi: 10.1038 / s41467-020-18058-8
15. Wec AZ, Wrapp D, Herbert AS, Maurer DP, Haslwanter D, Sakharkar M, et al. Amplia neutralización de los virus relacionados con el SRAS mediante anticuerpos monoclonales humanos. Science (2020) 369: 731–6. doi: 10.1126 / science.abc7424
PubMed Abstract | CrossRef Texto completo | Google Académico
16. Kubagawa H, Bertoli LF, Barton JC, Koopman WJ, Mestecky J, Cooper MD. Análisis del transporte de paraproteínas a la saliva mediante el uso de anticuerpos anti-idiotipo. J Immunol (1987) 138: 435-9.
17. Blanco-Melo D, Nilsson-Payant BE, Liu WC, Uhl S, Hoagland D, Moller R, et al. La respuesta desequilibrada del host al SARS-CoV-2 impulsa el desarrollo de COVID-19. Móvil (2020) 181 (5): 1036–45.e9. doi: 10.1016 / j.cell.2020.04.026
PubMed Abstract | CrossRef Texto completo | Google Académico
18. Vabret N, Britton GJ, Gruber C, Hegde S, Kim J, Kuksin M, et al. Inmunología de COVID-19: estado actual de la ciencia. Inmunidad (2020) 52: 910–41. doi: 10.1016 / j.immuni.2020.05.002
PubMed Abstract | CrossRef Texto completo | Google Académico
19. Brandtzaeg P. Inmunidad secretora con especial referencia a la cavidad bucal. J Oral Microbiol (2013) 5: 20401. doi: 10.3402 / jom.v5i0.20401
20. Mikhak Z, Agace WW, Lustre AD. Tráfico de linfocitos a los tejidos de las mucosas. En: Mestecky J, Strober W, Russell MW, Kelsall BL, Cheroutre H, Lambrecht BN, editores. Inmunología de las mucosas , 4. Amsterdam: Academic Press / Elsevier (2015). pag. 805-30.
21. Conley ME, Delacroix DL. Inmunoglobulina A intravascular y mucosa: ¿Dos sistemas de defensa inmunitaria separados pero relacionados? Ann Int Med (1987) 106: 892–9. doi: 10.7326 / 0003-4819-106-6-892
PubMed Abstract | CrossRef Texto completo | Google Académico
22. Russell MW, Kilian M, Mantis NJ, Corthésy B. Actividades biológicas de las inmunoglobulinas mucosas. En: Mestecky J, Strober W, Russell MW, Kelsall BL, Cheroutre H, Lambrecht BN, editores. Inmunología de las mucosas , 4. Amsterdam: Academic Press / Elsevier (2015). pag. 429–54.
23. Bidgood SR, Tam JC, McEwan WA, Mallery DL, James LC. La IgA translocalizada media la neutralización y estimula la inmunidad innata dentro de las células infectadas. Proc Natl Acad Sci USA (2014) 111 (37): 13463–8. doi: 10.1073 / pnas.1410980111
PubMed Abstract | CrossRef Texto completo | Google Académico
24. Griffiss JM, Goroff DK. La IgA bloquea la lisis inmunitaria iniciada por IgM e IgG mediante mecanismos moleculares separados. J Immunol (1983) 130: 2882-5.
25. Russell MW, Reinholdt J, Kilian M. Actividad antiinflamatoria de los anticuerpos IgA humanos y sus fragmentos Fabalpha: inhibición de la activación del complemento mediada por IgG. Eur J Immunol (1989) 19: 2243–9. doi: 10.1002 / eji.1830191210
PubMed Abstract | CrossRef Texto completo | Google Académico
26. Tomaras GD, Ferrari G, Shen X, Alam SM, Liao HX, Pollara J, et al. La IgA plasmática inducida por la vacuna específica para la región C1 de la envoltura del VIH-1 bloquea la unión y la función efectora de la IgG. Proc Natl Acad Sci USA (2013) 110: 9019–24. doi: 10.1073 / pnas.1301456110
PubMed Abstract | CrossRef Texto completo | Google Académico
27. Nurkic J, Numanovic F, Arnautalic L, Tihic N, Halilovic D, Jahic M. Significado diagnóstico de la IgA reducida en niños. Med Arch (2014) 68: 381–3. doi: 10.5455 / medarh.2014.68.381-383
PubMed Abstract | CrossRef Texto completo | Google Académico
28. Fischer A. Resistencia de los niños al Covid-19. ¿Cómo? Mucosal Immunol (2020) 13: 563-5. doi: 10.1038 / s41385-020-0303-9
PubMed Abstract | CrossRef Texto completo | Google Académico
29. Mestecky J, Hammarström L. Enfermedades asociadas a IgA: niveles reducidos de IgA – deficiencia de IgA. En: Kaetzel CS, editor. Mucosa Defensa inmunológica: Inmunoglobulina A . Nueva York: Springer (2007). pag. 330–86.
30. Moldoveanu Z, Clements ML, Prince SJ, Murphy BR, Mestecky J. Respuestas inmunitarias humanas a las vacunas contra el virus de la influenza administradas por vías sistémicas o mucosas. Vaccine (1995) 13: 1006–12. doi: 10.1016 / 0264-410x (95) 00016-t
PubMed Abstract | CrossRef Texto completo | Google Académico
31. Mestecky J, Jackson S, Moldoveanu Z, Nesbit LR, Kulhavy R, Prince SJ, et al. Escasez de respuestas de IgA específicas de antígeno en sueros y secreciones externas de individuos infectados por VIH de tipo 1. AIDS Res Hum Retrovirus (2004) 20: 972–88. doi: 10.1089 / aid.2004.20.972
PubMed Abstract | CrossRef Texto completo | Google Académico
32. Zhang X, Tan Y, Ling Y, Lu G, Liu F, Yi Z, et al. Factores virales y del huésped relacionados con el resultado clínico de COVID-19. Nature (2020) 583: 437–40. doi: 10.1038 / s41586-020-2355-0
PubMed Abstract | CrossRef Texto completo | Google Académico
33. Jackson S, Mestecky J, Moldoveanu Z, Spearman P. Apéndice I: Recolección y procesamiento de secreciones mucosas humanas. En: Mestecky J, Bienenstock J, Lamm ME, Mayer L, McGhee JR, Strober W, editores. Inmunología de las mucosas , 3. Amsterdam: Elsevier / Academic Press (2005). pag. 1829-1839.
34. Callow KA. Efecto de la inmunidad humoral específica y algunos factores inespecíficos sobre la resistencia de los voluntarios a la infección respiratoria por coronavirus. J Hyg (Londres) (1985) 95: 173–89. doi: 10.1017 / s0022172400062410
35. Isho B, Abe KT, Zuo M, Jamal AJ, Rathod B, Wang JH y col. Persistencia de las respuestas de anticuerpos en suero y saliva a los antígenos pico del SARS-CoV-2 en pacientes con COVID-19. Sci Immunol (2020) 5: eabe5511. doi: 10.1126 / sciimmunol.abe5511
PubMed Abstract | CrossRef Texto completo | Google Académico
36. Cervia C, Nilsson J, Zurbuchen Y, Valaperti A, Schreiner J, Wolfensberger A, et al. Secreción de anticuerpos sistémicos y mucosos específicos de SARS-CoV-2 durante COVID-19 leve versus grave. bioRxiv (2020) 2020.05.21.108308. doi: 10.1101 / 2020.05.21.108308
37. Sterlin D, Mathian A, Miyara M, Mohr A, Anna F, Claer L, et al. La IgA domina la respuesta temprana de anticuerpos neutralizantes al SARS-CoV-2. medRxiv (2020), 2020.06.10.20126532. doi: 10.1101 / 2020.06.10.20126532
38. Russell MW, Mestecky J. Vacunas mucosas: descripción general. En: Mestecky J, Strober W, Russell MW, Kelsall BL, Cheroutre H, Lambrecht BN, editores. Inmunología de las mucosas , 4. Amsterdam: Academic Press / Elsevier (2015). pag. 1039–46.